
Landscape ecology is the study of the relationships between landscape systems and their many ecological functions and processes. Landscapes can be functional or dysfunctional with respect to native* and exotic (invasive) vegetation components, the creation and expansion of novel ecosystems, and the multitude of human activities and their effects upon the landscapes. Some of the best research available on the subject, including concepts and monitoring models, is the work of John Ludwig and David Tongway. Their publications are numerous, but most useful is, “Landscape Ecology, Function and Management: Principles from Australia’s Rangelands” (1997).
Their book is the product of decades of experience in the semi-arid and arid lands of Australia, and brings forth many important and grounded concepts about restoration, overgrazing, and landscape functionality. Though the theories and conclusions were largely developed Down Under, the principles are readily applied to North American drylands of the same aridity and elsewhere. Why this book has remained below the radar of many land managers is beyond me. It is an exceptional landscape ecology text, with easy to read prose and basic but effectively supportive photographs illustrating the various concepts presented. Unfortunately, little to none of their works are widely taught in schools.
The book explains the consequences of overgrazing, various landscape function and dysfunction indicators, and provides a look at rangeland management at the landscape scale, which is sorely missing from today’s major rangeland management activities; which are still more or less focused on growing more forage to graze more cattle. Taken together, these aspects present the big picture, the one that fundamentally matters where ecology is concerned. By integrating practical landscape-scale concepts into land management theories, especially those involving rangeland management and rangeland restoration, the book is a far superior resource than what has been published over the last 20 years. “Landscape Ecology, Function and Management: Principles from Australia’s Rangelands” is a prominent publication and a wise investment of one’s reading time and future referrals.
Too often as land managers, we sometimes get tunnel vision and become engrossed in the small things, such as how many species are in restoration seed mixes, species richness in a restoration planting, biodiversity or even the out-of-reach pre-settlement plant community composition. While those elements are important, they can shift our focus away from the concepts that really matter, causing us to miss out on big picture truths. West (1983) captures this simply, “It is more important to preserve [ecosystem] processes than all organisms, when functional redundancy can be demonstrated.” Species richness in restoration plantings is always nice to see and helps to recruit public support, but if we focus too much on generating diversity and richness over conservation of foundation species and critical landscape functions, we lose ground over time to exotic species and loss of system functions that generate goods and services to society, and help to hold a degrading world together.
Unfortunately today, many remnant native ecosystems have suffered some form of degradation, even those few special remnants ecologists have defined as being in great shape. Most systems have crossed an ecological threshold and are beyond the reach of today’s restoration capabilities. As a definition, a threshold shows the following: a boundary across spatial (space) and temporal (time) scales, by which the shift across the threshold is not readily or easily reversible to the previous state. That is, not without intensive mechanical and chemical work, and regular inputs to maintain the “restored” state (Friedel 1991). Knowing whether a particular landscape is demonstrating a functional or dysfunctional state can help restoration ecologists determine whether or not a particular landscape is suitable for restoration work. Landscape degradation and dysfunction are the dynamics of the overall process of desertification, an issue of ancient origins and global consequences (Whitford 2002, Verstraete 1986).

Map Source: http://www.fao.org/docrep/007/y5738e/y5738e04.jpg Drylands occupy some 45% of the globe, home to 20% of the global human population. What happens in drylands affects the rest of the world, as the earth is a dynamic and contiguous system. The more research we have on dryland systems and their functional and dysfunctional dynamics, the more educated our decision making can be at the global, state, and local policy levels.
The basic and most common visual definition of an ecological threshold shift is the conversion of desert grassland to shrubland. The bulk of this shift occurred most rapidly and extensively over the last 150 years, largely due to past and current chronic overgrazing in combination with other environmental factors (Van Auken 2000), notably the relatively steady increase in atmospheric carbon dioxide (CO2), which probably facilitates increases in woody species by virtue of their C3 photosynthetic pathway (Polley 1997). Other factors are at play as well and are thoroughly discussed in D’Odorico et al. (2012). Moving forward from the end of the grassland system’s reign is more than likely a hybrid (novel + native) system of native woody species and increased richness in exotic herbaceous species, many of which will become naturalized (Archer and Predick 2008).
For now, we can study remnant grasslands and observe and record what the causes are for their declines. As an example, a key indicator of landscape dysfunction within the grassland biomes of the world is the loss of perennial grass components – the keystone foundation of most grasslands. As perennial grass communities exhibit lowered performance, become impaired or locally extinct due to past land use legacies, annual forb communities increase in abundance and become established. This dynamic is particularly strong and entrenched in arid grasslands of the Desert Southwest, especially during and after wet winters, although it is no less a contribution to dysfunction in semi-arid grasslands (“drylands” collectively). The causes of this dysfunction are many, but chief among them is overgrazing, beginning in the 1600s (Ford 1987), with major peaks in the 1890s, another during WWI, and during the 1920s, just prior to the Dust Bowl years (Havstad et al., 2006). Overgrazing is a damaging process that is highly variable throughout time and space, is inherent to the livestock industry (Freudenberger et al. 1997), and intensifies the pace of degradation.

Note the remaining perennial grassy small-scale patches and some of the shrub bases show the pedestal effect as a result of sheet erosion. The dominant grass species here is the exotic invasive Lehmann Lovegrass (Eragrostis lehmanniana). The majority of the Desert Southwest appears as pictured.
Above, this former desert grassland is now mostly Creosotebush (Larrea tridentata) and Mesquite (Prosopis spp.) shrubland with large bare ground interpsaces. This site is being increasingly controlled by abiotic factors (geomorphology) rather than biotic factors (plant-soil interactions). The bare areas allow increased rates of wind and water erosion, and increased ultraviolet radiation, which causes changes in soil propertiest, furthering the loss and subsequent local extinction of native perennial grassland components. Such a process has occurred extensively across vast areas of the Desert Southwest and the American West.

Areas controlled largely or wholly by abiotic factors, such as the site pictured here, lack any geomorphic stability and typically are not candidates for grassland restoration, as the cost of such work is far too great for such minimal to no returns. Additionally, a lack of ecotypically appropriate seed sources available on the commercial market, excepting ‘Van Horn’ Green Sprangletop, exist for this site (Chihuahuan Desert, Trans-Pecos, TX). Research and development in Texas to increase commercial availability of ecotypically appropriate certified native seed sources is ongoing (Smith et al. 2013).
Further, in desert grasslands of the Southwest, woody and semi-woody plants population expansion via encroachment (“invasion” by other authorities) rarely give up their newly claimed lands (Ellison 1960, Wright and Honea 1986), even with livestock grazing removed from sites. The persistence of woody species and their creation of steady states (systems that show greater stability than previous systems) is often in conjunction with exotic herbaceous species (i.e. Mesquite and Lehmann Lovegrass on sandy sites). This is a foundational degradation dynamic in arid and semi-arid grasslands. On a positive note, the grassland-shrubland transition shows great promise as a superior carbon sink compared to remnant or restored grasslands, especially during drier climatic times (Petrie et al. 2015).

The dominant indicator of landscape dysfunction caused by overgrazing here is Perennial Broomweed (Gutierrezia sarothrae). Note the stunted perennial grass remnants (mostly Tobosagrass and some Vine Mesquite) barely offering cover across the interspaces between the indicator forb and the shrubs. The shrubs are Mesquite, Tarbush and Creosotebush.
As a whole, landscape dysfunction is a multivariate process and the results (loss of functions) are neither bad nor good, they just are. Dysfunction can progress slowly or rapidly, evidently or obscurely. Should overgrazing or other exploitation continue, dysfunction will, with increasing system losses, progress to degradation, and further, to desertification. Landscape degradation is a hallmark of global human occupation (Vitousek et al. 1997). We can either hide from the consequences and do nothing, engage in illogical thinking (“restore to pre-European conditions!”, “Saving the great grasslands of the world with more cattle grazing!”), or we can accept the damages of our fascinating modern progress, learn the many dynamic intricacies involved, and where applicable, apply the lessons learned as true restoration practices.
How we perceive landscape changes and subsequently classify those changes depends greatly on how we value landscapes. Some of us value landscapes solely for their use as areas of livestock production, however short-term and narrowly-focused that use is. Others view landscapes as a place to let the system do its thing, to observe its changes and interactions with a multitude of environmental factors. Such a broader view allows for a great deal of science to be practiced and knowledge to be gained. The way in which a person views their world through a social lens and the culture through which they were raised greatly impacts the formation of the values they place on landscapes (Tongway and Ludwig 1997). Aldo Leopold’s formation of his famous land ethic was decades in the making, and only after he committed many environmental mistakes; the most famous of which was his killing a wolf and later realization: “I was young then, and full of trigger-itch; I thought that because fewer wolves meant more deer, that no wolves would mean hunters’ paradise. But after seeing the green fire die, I sensed that neither the wolf nor the mountain agreed with such a view” (Leopold 1949).
In order to keep up with the changing values of our anthropocentric world, current ecological views and paradigms must shift to actively include broader landscape-scale dynamic thinking, while simultaneously increasing our ability to restore appropriate landscapes to the degree that such lands will continue to provide the goods and services the modern world has come to depend on. The gap between functional and dysfunctional landscapes continues to widen in correlation with increased human activities, needs and current climatic changes. Happy mediums need to be met between needs of producers and needs of landscapes, and the increasing constraints of pastoralism, especially in drylands, needs to be realized and appropriately integrated at various policy levels.
The basic indicators of landscape dysfunction as observed by others and myself in drylands is illustrated below.
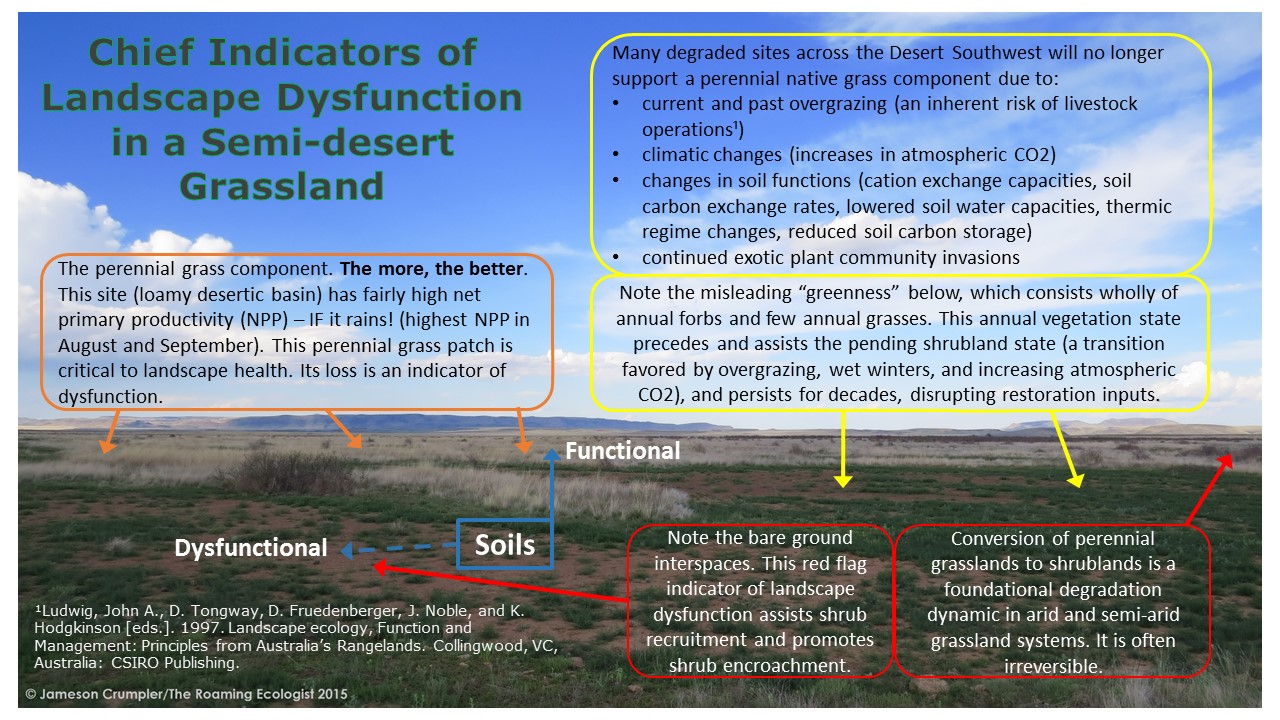 *Native here means species that existed on the North American continent prior to arrival of Europeans. It’s a relative term.
*Native here means species that existed on the North American continent prior to arrival of Europeans. It’s a relative term.
References
Archer, S.R. and Predick K.I. 2008. Climate change and ecosystems of the southwestern United States. Rangelands. 30(3):23-28.
Ellison, L. 1960. Influence of grazing on plant succession of rangelands. The Botanical Review. 26(1): 1-78.
Ford, Richard I. 1987. The new Pueblo economy. In: Ellis H. Florence, M.E. Jenkins, and F.I. Ford [eds.]. 1987. When cultures meet: Remembering San Gabriel del Yunge Oweenge, pp. 73-87. Sunstone Press. Santa Fe, NM, USA.
Freudenberger, D., K. Hodgkinson, and J. Noble. 1997. Causes and consequences of landscape dysfunction in rangelands. In: Ludwig, John A., D. Tongway, D. Fruedenberger, J. Noble, and K. Hodgkinson [eds.]. 1997. Landscape ecology, function and management. Collingwood, VC, Australia: CSIRO Publishing. p. 63-77.
Friedel, M.H. 1991. Range condition assessments and the concept of thresholds: A viewpoint. Journal of Range Management. 44(5):422-426.
Havstad, Kris M., L.F. Huenneke, W.H. Schlesinger. [eds.]. 2006. Structure and function of a Chihuahuan Desert ecosystem. New York City, New York, USA: Oxford University Press. 492p.
Leopold, Aldo. 1949. A Sand County Almanac. New York City, New York, USA: Oxford University Press. 226p.
Petrie, M.D., S.L Collins, A.M. Swann, P.L. Ford and M.E. Litvak. 2015. Grassland to shrubland state transitions enhance carbon sequestration in the northern Chihuahuan Desert. Global Change Biology. 21: 1226–1235.
Polley H.W. 1997. Implications of rising atmospheric carbon dioxide concentration for rangelands. Journal of Range Management. 50:562–577.
Smith, F.S., K.A. Pawelek, A.D. Falk, J. Lloyd-Reilley, J.P. Muir, J. Breeden, B.J. Warnock, C.S. Shackelford, and M.A. McCraw. South Texas Natives and Texas Native Seeds Projects. In: Proceedings of the 67th Southern Pasture and Forage Improvement Conference. 2010 Apr 22-24; Tyler, TX: Texas A&M AgriLife Extension.
Tongway, D. and Ludwig, J. 1997. Nature of landscape dysfunction. In: Ludwig, John A., D. Tongway, D. Fruedenberger, J. Noble, and K. Hodgkinson [eds.]. 1997. Landscape ecology, function and management. Collingwood, VC, Australia: CSIRO Publishing. p. 49-61.
Van Auken, O.W. 2000. Shrub invasions of North American semiarid grasslands. Annual Review of Ecology and Systematics. 31:197-215.
Verstraete, M.M. 1986. Defining desertification: A review. Climatic Change. 9:5-18.
Vitousek, P.M., H.A. Mooney, J. Lubchenco, and J.M. Melillo. 1997. Human domination of Earth’s ecosystems. Science. 227:494-499.
West, N.E. 1993. Biodiversity of rangelands. Journal of Range Management. 46:2−13.
Whitford, Walt G. 2002. Desertification. In: Whitford, Walt G. Ecology of desert systems. 2002. San Diego, CA, USA: Academic Press. p.275-304.
Wright, R.A., and J.H. Honea. 1986. Aspects of desertification in southern New Mexico, USA: Soil properties of a mesquite duneland and a former grassland. Journal of Arid Environments. 11:139-145.


{kind=link}